
Tobacco Hornworm, Manduca sexta in larvel, pupal, and adult stages of development
Tobacco Hornworm, Manduca sexta in larvel, pupal, and adult stages
of development
Manduca sexta-sexta meaning "six-fold", Manduca, the Latin word for "glutton." These moths have six pairs of yellow spots on their abdomens and while they are caterpillars, consume an average of 201 square inches of leaf between their fourth and fifth molts. Although a menace to tobacco growers, hornorms are easily reared in the laboratory on artificial diets and are large enough that surgery on their nervous system is not extremely difficult. Due to their latter attributes, hornworms have been viewed as a particularly appealing system to researchers interested in neuroethological topics such as ecdysis behavior. Ecdysis is the insect molting process where by the outer cuticle is shed and the insect emerges into a more mature stage of its life.
Probably the most conspicuous features of arthropods is the periodic shedding of the cuticle. The act of molting or ecdysis punctuates cylces of growth and development. In a developing fifth-instar larva Manduca Sexta, a repetative sequence of muscular contractions can be seen along the entire length of its abdomen (abdominal and thoracic segments). This lasts from 60 to 80 min, occurs approximately 1.5 hours prior to ecdysis and is known as pre-ecdysis behavior. These contractions begin in the most posterior segment and gradually spread anteriorly. Each cycle begins with synchronous contractions of all abdominal segments on both sides of the animal's dorsal midline and last for about one second. This is followed by contraction of lateral muscles above the abdominal prolegs on one side of the body, which is in turn followed by the contraction of the same set of muscles on the other side of the body. These movements serve to loosen the old cuticle and continue until ecdysis behavior is initiated.
Ecdysis behavior is characterised by a distinctly different motor pattern than that which is demonstrated during pre-ecdysis behavior. Ecdysis lasts for approximately 10 minutes and consists of peristaltic waves of contractions where each segment of the animal is lifted and contracted forward in a posterior to anterior sequence. As the forlegs and thoracic legs are freed from the old cuticle, a split develops behind the old head capsule which allows the larva to emerge.
Each larval molt in Manduca Sexta consists of a similar series of behavioral events and developmental markers used to stage larvae during the molt. What differs between the cylces of molts is the timing of the sequence of events such that the events associated with earlier molts are proportionately compressed. Thus, although the above description of behavioral events is directly associated only with the fifth instar molt, the same pattern is found in other molts.
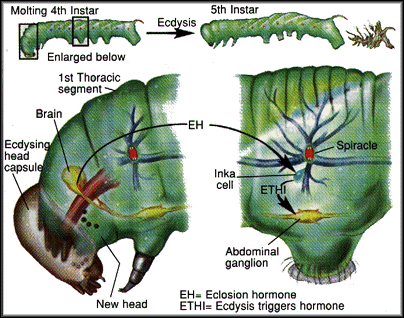
Mechanism of ecdysis control
Recent findings suggest that ecdysis in Manduca Sexta is mediated by eclosion hormone (EH) and an ecdysis triggering hormone (Mas-ETH or ETHI). Previously it was believed that EH, a 62 amino acid peptide released from the brain, was the sole mediator of preecdysis and ecdysis behaviors. However, Mas-ETH has been implicated in the timing of ecdysis. Successful ecdysis requires withdrawal of the tracheal linings (important for oxygen delivery) from the old cuticle. It appears that EH stimulates Inka cells to release Mas-ETH, a 26 amino acid peptide, which in turn initiate the abdominal contractions by stimulating the abdominal ganglion. However, ecdysis is fatal to the animal unless the Inka cells know when to stimulate the abdominal ganglion. It appears that the Inka cells monitor tracheal linings as they pass from the old cuticle through the spiracle to the new cuticle. Only after the fragile trachea is withdrawn is the animal ready to commit itself to molting.
Diagram and explanation modified from Truman (1996).
Early classics:
Truman, JW (1978). Hormonal release of stereotyped
motor programmes from the isolated nervous system of the cecropia silkmoth.
Journal of Experimental Biology 74: 151-173.
Copenhaver, PF and Truman, JW (1982). The role of eclosion hormone in the larval ecdyses of manduca sexta. Journal of Insect Physiology 28: 695-701.
Reviews:
Evans, HE (1985). The Tobacco Hornworm. In: The
Pleasures of Entomology, Smithsonian Instiution, pp. 145-156.
Reynolds, SE (1980). Integration of Behavior and Physiology in Ecdysis. In: Advances in Insect Physiology, Berridge MJ, Treherne JE, & Wigglesworth VB, eds., pp. 475-579.
Truman, J.W. (1992) Hormonal regulation of behavior: insights from invertebrate systems. In J.B. Becker, S.M. Breedlove, and D. Crews (Eds.), Behavioral Endocrinology. Cambridge, MA: MIT Press. Pp. 423-450.
Some Recent Advances:
Ewer J, De Vente J, & Truman JW (1994). Neuropeptide
induction of cyclic GMP increases in the insect CNS: Resolution at the level
of single identifiable neurons. Journal of Neuroscience 14: 7704-7712.
Truman JW (1996). Ecdysis control sheds another layer. Science 271: 40-41.
Zitnan D, Kingan TG, Hermesman JL, & Adams ME (1996). Identification of Ecdysis-triggering hormone from an epitracheal endocrine system. Science 271: 88-91
