Figure 1: From: http://www.unc.edu/depts/oceanweb/turtles/
True navigation is when an animal returns to a place without using landmarks (cues) from the destination and the outward journey (Boles and Lohmann, 2003). You may wonder how that’s possible considering how the use of such information is a typical way of navigating. Just think of how you find your way to the grocery store. You probably rely on many visual landmarks, such as passing a church, a gas station or a school. If you’ve traveled the route many times, you may even be able to do it blindfolded by using non-visual landmarks, such as odor, sound and proprioception (e.g.: feeling your body move forward a given distance and knowing when to turn). Studies have shown that in addition to using those types of landmarks, some animals use the earth’s magnetic field to navigate. In other words, some animals have an internal compass that enables them to sense direction. This ability has also been likened to a global positioning system (GPS).
Below is an overview of:
1. what magnetic information is and how different species use it to navigateResources and references on magnetic information are listed toward the bottom of the page.
2. model systems that are used to study how magnetic information is used
3. theories of how the brain processes magnetic information
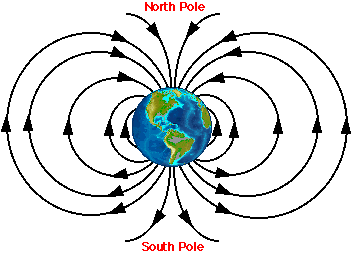
The liquid core of the Earth gives off a magnetic field. Energy goes from the Earth’s South Pole and reenters the Earth at the North Pole (see Figure 1, taken from the Lohmann Lab website). Evidence suggests that animals can navigate by detecting the strength of the magnetic field and the angle at which the field meets the earth (Lohmann and Lohmann, 1996, Lohmann and Lohmann 1994, Light and Salmon, 1993), which are distinct for each spot on the globe.
Figure 1: From:
http://www.unc.edu/depts/oceanweb/turtles/
Many animals respond to the Earth’s magnetic field and can use it as a navigation tool. Here are descriptions of how a few of those species use magnetic information.

From:
http://www.angelfire.com/ga/huntleyloft/Page1.html
Beginning at about 6 weeks of age, pigeons adopt a home loft that remains permanent throughout their lives. Pigeons use different landmarks to return to the home loft, including visual and olfactory (smell) landmarks. Pigeons also use magnetic information to navigate. When the magnetic field is disrupted by putting magnetic coils around their heads (Walcott, 1979) or by gluing magnets to the back of their necks (Keeton (1971)), pigeons cannot return to their loft. Navigation is also disrupted when pigeons must journey through anomalies, which are naturally occurring magnetic disruptions ( Wiltschko and Wiltschko, 1996).

From:
http://www.unc.edu/depts/oceanweb/turtles/
Florida beaches are a common hatching area for loggerhead sea turtles. After the turtles hatch, they crawl across the beach to the ocean and then out to sea. Examples of different swim routes that individual turtles take are shown in Figure 2.
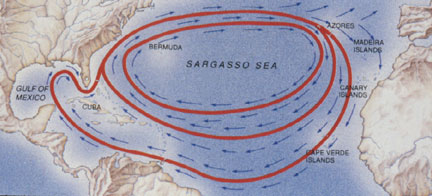
Figure 2: From:
http://www.unc.edu/depts/oceanweb/turtles/
Hatchlings swim along the east coast of the United States and then swim northeast with the Gulf Stream. Female turtles remain in the North Atlantic for a few years. They complete the circle within the Atlantic Ocean (see Figure 2), and then they return to their natal beach to nest. The return trip can take them across hundreds or thousands of km of open ocean.
Data from the Lohmann lab indicate that turtles use various cues to orient themselves:
1. from the natal nest toward the shore,
2. to the open sea, and
3. along their migratory route.
It is thought that hatchlings use visual cues to orient themselves toward the shore (where there is more light) and then use the direction of the waves to orient themselves out to sea. Magnetic information becomes important once they are in the open sea. ( Lohmann and Lohmann, 1996). This can be studied in the laboratory by observing the swim direction of hatchlings in a small pool. When the magnetic field surrounding the swimming hatchlings is reversed, the hatchlings change their swim direction such that it matches the swim direction of their migratory path. More recently it was shown that when hatchlings are exposed to magnetic fields that mimic different natal beaches they redirect their swim direction to maintain the same migratory path ( Lohmann et al., 2001). See the Lohmann lab website for more information: http://www.unc.edu/depts/geomag/

From:
http://www.unc.edu/depts/oceanweb/lobsters/bigspinylobster.jpg
Spiny lobsters live in caves, come out to forage for food and then return to the same cave. They also have an annual migration. Experimentally, they have been shown to return to their cave after being placed in an unfamiliar location. Further, lobsters that were exposed to alterations in the magnetic field oriented themselves back toward their caves. (Lohmann et al., 1995, Boles & Lohmann, 2003).
Bacteria
Honeybees
Mosquitoes
Butterflies
Some mollusks
Newts
Rainbow trout
Salmon
Dolphins
Mole rats
Evidence for and against magnetic orientation in humans is limited. The discovery of magnetic particles (magnetite) in the human brain (Dunn et al., 1995) suggests that humans may be sensitive to magnetic fields (see next section on how the brain processes magnetic information). Some studies reported that humans can point to the North when they have limited other cues ( Baker, 1980), while other studies (e.g.: Fildes et al., 1984) found no evidence of magnetic orientation by humans. Regardless of whether magnetic orientation exists in humans, humans definitely prefer to use visual cues for navigation.
How the brain processes magnetic information remains unclear. Some species have particular brain areas
that respond to magnetic information, and there may be nerve cells in those areas that detect changes
in the magnetic field. In a theoretical paper, Walker et al. (2002)
proposed that
animals may be able to detect magnetic field information due to specialized nerve cells. Similar to
how we have specialized nerve cells for senses such as vision and olfaction, Walker et al. hypothesized
that there are nerve cells that react to different characteristics of the
magnetic field such that the animal can respond behaviorally. Such cells are called “magnetoreceptors”
(mag-NEE-toe-ree-sep-tors). We can think of them as an internal compass that is hooked up to the body
such that the “needle” of the compass moves with the changing magnetic field. When the needle is
displaced, it can trigger the activation of other cells that help the animal sense where to direct
its movements.
It is believed that the presence of the naturally magnetic mineral magnetite (a.k.a. Fe3O4 or lodestone)
in the brains of some animals is one way that magnetoreception works (
Kirschvink et al., 2001).
Studies from rainbow trout showed that brain areas that are sensitive to the magnetic field have nerve
cells that contain magnetite (Walker et al., 1997,
Diebel et al., 2000). Currently, the neural mechanisms
that underlie magnetoreception are largely unknown.
Earth’s Magnetic Field:
http://csep10.phys.utk.edu/astr161/lect/earth/magnetic.html
http://liftoff.msfc.nasa.gov/academy/space/mag_field.html
Magnetoreception:
http://www.animalbehavioronline.com/magnetoperception.html
http://www.ks.uiuc.edu/Research/magsense/
http://www.wowessays.com/dbase/af2/rsk183.shtml
Orientation in Pigeons:
http://www.albertaclassic.com/suncomp.php
http://www.animalbehavioronline.com/pigeonhoming.html
Orientation in Sea Turtles:
http://www.unc.edu/depts/oceanweb/turtles/
Orientation in Lobsters:
http://www.unc.edu/depts/oceanweb/lobsters/lobintro.html
Blakemore, R. (1975) Magnetotactic bacteria. Science 190, 377-379.
Keeton, W.T. (1971) Magnets interfere with pigeon homing. Proc Nat Acad Sci 68, 102-106.
Walcott, C. (1979) Pigeons have magnets. Science 205, 1027-1029.
Kirschvink, J.L., Jones, D.S. & MacFadden, B.J. (eds.) Magnetite biomineralization and magnetoreception in organisms: A new biomagnetism. New York: Plenum Press, 1985.
Wiltschko, W., Wiltschko, R. (1996) Magnetic orientation in birds. J Exp Biol 199, 29-38.

{kind=link}