
A female (left) and male (right) zebra finch, Taeniopygia guttata
A female (left) and male (right) zebra finch, Taeniopygia guttata
The learning of birdsong has become one of the most popular model systems in the neuroethology community for understanding how experience modifies the brain, and interacts with non-experiential aspects. All oscine songbirds will develop highly degraded song if they are deafened during development, and develop degraded but species-typical songs if reared in isolation. Sub-oscines develop normal song even with deafening; their song is not learned but rather genetically programmed. The oscine zebra finch has become especially popular in neurobiology laboratories, partially due to the ease with which they are maintained and bred in captivity (without regard to season), rapid sexual maturity (90 days after hatching), and the fact that the phenomena under study, the learning of song, is a robust behavior that follows a stereotypic pattern following hatching, making it amenable to genetic approaches as well as standard neuroanatomical, neuropharmacological, and electrophysiological methods. These are sexually dimorphic animals: only the males sing, the plumage of males has various markings that the females lack, and males have red beaks and cheek patches while females have orange beaks. Sexual selection is done by the female. They are native to the semi-arid regions of Australia and Timor, nesting in large social groups of 20 to 1000 birds.
Tree of Life: Passeriformes (family Passeridae, sub-family Estridinae, tribe Poephilini, genus Taeniopygia)
The learning of birdsong by developing young is a process that begins with the production of subsong vocalizations and ends with the production of a rendition of the tutor's song (usually, the father). The acquisition of birdsong has a "sensitive period" in some species, including zebra finches, where interfering with the development of song during this period results in permanent impairment. In zebra finches, young will form an auditory model of the tutor's song as early as 20 days after hatching, and song acquisition will be completed by around 35 days. The song becomes consistent through rehersal by around 60 days, and crystallizes (becomes invarient) at sexual maturity, approximately 90 days. Although the development of bird song has a relatively clear and stereotypic pattern, birdsong's precise role in their subsequent day-to-day activities is still an area of research.
Obtaining and retaining territory, and attracting and stimulating females are some of the leading candidates for the roles of birdsong, not surprisingly given the predominance of these behaviors in the life of a bird. Many open questions exist, such as the relation between fitness and the properties of the male's song (if any) in the female's sexual selection process, and the role of other (non-song) vocalizations in these behaviors (Kroodsma and Byers, 1991).
There is evidence that unpaired males sing more than paired males; in some species the male will stop singing, or shift the use of song once a female is near. Again, the role of other vocalizations has not been ascertained. One of the few exceptions is one study where a special surgery preserved non-song vocalizations while eliminating song (McDonald 1989). This study found the male's ability to obtain and maintain territory was clearly decreased. A currently popular evolutionary scenario takes song to be a selected trait whose expression is dependent on the condition of the male. The female can then effectively listen for aspects of song (ranging from size of repertoire to song loudness, which can be correlated to the size of the bird's territory and thus to their ability to support young via the energetic cost of singing), which have been tied to subsequent clutch fitness in the ancestral condition. There are some hypotheses as to why learned birdsong, as opposed to genetically programmed birdsong, evolved: 1) to allow for more complex song via cultural transmission rather than genetic transmission; 2) learned song allows for the development of intra-species varients of song, called dialects, which may serve to segregate populations of the same species; 3) learned song allows for more rapid adaptation of the song to different acoustic environments (Slater, 1989)
Here's a sample zebra finch song and description of some of its acoustic features. Clicking on the sonogram will play the song.
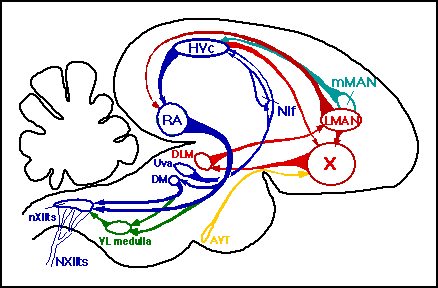
A schematic of a male bird's brain and song circuit. This is a cross-sectional
view through the center of the brain, where the back of the head would be
to the left and the beak to the right. Diagram courtesy Heather
Williams.
Abbreviations: AVT: Area Ventralis of Tsai DLM: Dorso-Lateral division of the Medial thalamus DM: Dorso-Medial subdivision of the intercollicular nucleus HVc: High Vocal Center (aka Hyperstriatum Ventralis pars caudalis) LMAN: Lateral Magnocellular nucleus of the Anterior Neostriatum (aka IMAN) MMAN: Medial nucleus Magnocelularis of the Anterior Neostriatum NCM: Neostriatum Caudo-Medial nucleus NIf: interfacial nucleus of the neostriatum NXIIts: tracheosyringeal portion of the 12th cranial nerve (tracheosyringeal nerve) nXII: nucleus of the 12th cranial nerve (hypoglossal) RA: Robust nucleus of the Archistriatum Uva: nucleus Uvaeformis of the thalamus VL medulla: VentroLateral medulla X: Area X of the paraolfactory lobe.
Color key and circuit summary:
Blue (production pathway) is the song output pathway, starting at Uva and terminating at the syrinx (not shown), which is the bird's vocal organ. Disturbances to this pathway affects song, regardless of whether the disturbance occurs after or before the learning of song.
Red (learning pathway) is the pathway that connects HVC to RA via area X, DLM, and LMAN. Since collaterals of neurons in LMAN project to area X, this forms a recursive loop. Disturbances to this pathway will affect song development, but not the production of song in the adult male.
Turquoise is MMAN, a nucleus that sends its output to HVC, and Yellow is AVT, which supplies dopamine to area X, and Green is a hindbrain structure that has descending projections to the nXIIts. The role of these structures in birdsong is an area of current research.
As the focus of this web page is the learning of song as distinct from the production of song, this summary will focus on the anatomical substrates of song development. The lateral magnocellular nucleus of the anterior neostriatum, LMAN, is the nucleus for which the evidence of a specific role in song development is clearest. If this area is lesioned from 20 to before 60 days in the developing male zebra finch, the bird will develop an abnormal song. Lesions in this same area in the adult (beyond 60 days) has little consequence for an already learned song, and no observable consequence when performed beyond sexual maturity at 90 days. As the circuit above shows, LMAN sends its only efferent to the robust nucleus of the archistriatum (RA), the primary song output nucleus. Based on several lines of evidence, it has been suggested that the LMAN projection to RA is the mechanism for the tutor song to selectively entrain the activity in the HVC-RA pathway so that vocal output is brought into correspondence with the tutor song. Recently, an interesting pattern of gene expression for a newly discovered protein, synelfin, in song nuclei was discovered. The most distinct pattern is in LMAN, where it was measured to vary from 1 1/4 times the level outside the nucleus to 3/4 of the level outside the nucleus after sexual maturity, most of this change occurring in the time before the acquisition of song at 35 days (George et al. 1995). Recent work by Yu and Margoliash (1996) suggests that HVC is responsible for the correct sequence of syllables in a song, while RA ensures the proper sequencing of subsyllabic structure, the notes within a syllable. One interesting implication of this work is that isolates, whose LMAN-RA projection is present but functionally useless given the above model, may have less distortion of syllable structure than of note structure compared to normals, presuming there is a higher level of commonality between conspecifics in syllable as opposed to note structure due to innate motor song production patterns.
Hormonal control of behavior: Cells in the LMAN, HVC, RA, and nXIIts accumulate testosterone or its metabolites, and there is some evidence that song production and learning may be androgen-dependent behaviors.
Early classics:
Nottebohm F, Stokes T, Leonard CM (1976) Central control of song in the canary. J Comp Neurol 165:457-486.
Bottjer SW, Miesner EA, Arnold AP (1984) Forebrain lesions disrupt development but not maintenance of song in passerine birds. Science 224:901-903.
Reviews:
Konishi, M. (1989). Birdsong for Neurobiologists. Neuron , 3, 541-549.
Kroodsma DE, Byers BE (1991) The function(s) of bird song. Am Zool
31:318-328.
Slater, P. J. B., Eales, L. A. and Clayton, N. S., (1988) Song learning in zebra finches (Taeniopygia guttata): progress and prospects. In: Advances in the Study of Behavior. 18. (Rosenblatt, J. S., Beer, C., Busnel, M.-C. & Slater, P.J. B., eds.) Harcourt
Slater, P. (1989). Bird song learning: causes and consequences. Ethology, Ecology, and Evolution, 1:19-46.
Some Recent Advances:
Fletcher, N.H. (1988). Bird song- A quantitative acoustic model. J. Theoretical Biol 134(4), 455-481.
George JM, Jin H, Woods WS, Clayton DF (1995) Characterization of a novel protein regulated during the critical period for song learning in the zebra finch. Neuron 15:361-372.
Mello CV, Nottebohm F, Clayton DF (1995) Repeated exposure to one song leads to a rapid and persistent decline in an immediate early gene's response to that song in zebra finch telencephalon. J Neurosci 15:6919-6925.
McDonald, M.V. (1989). Function of song in Scott's seaside sparrow, Ammodramus maritimus peninsulae. Anim. Behav. 38:468-485.
Yu, A.C., Margoliash, D. (1996) Temporal hierarchical control of singing in birds. Science 273: 1871-1875.
A web site on the theory of costly signalling by Carl Bergstrom
NEUROBIOLOGY OF BIRDSONG ---comprehensive list of links to birdsong neurobiology labs
David Clayton's Lab at Beckman/University of Illinois
The laboratory of Heather Williams
FinchWorld (oriented to aviculture)
BIRDNET American ornithological council web site-loads of info to satisfy your inner bird
