
Honey Bees, Apis mellifera.
Honey Bees, Apis mellifera.
Honeybees (Apis mellifera) live in colonies of thousands of individual bees working cooperatively to maintain the integrity of the nest and to ensure the survival of the next generation of bees. In order to accomplish these goals, honeybees have developed elaborate social structures to divide the many tasks among the worker bees.
Tree of Life: Apoidea
The many tasks of associated with maintaining the hive are divided among
the worker bees in an age-based fashion (polyethism). That is, individual
bees perform the tasks in a specific sequence. The youngest bees clean the
cells for about 3 days, then they are nurses to the brood for several days.
From days 10 to 20, the bees engage in building activities and pollen storage
and reception. At around day 20, the bees stand guard at the hive entrance,
and after a few days the bees become foragers. The bees will remain foragers
until they die. This pattern, however, is flexible. Young bees raised in
the absence of older foragers will begin to forage precociously. The mechanism
underlying this age-based division of labor and its ability to respond to
environmental changes is currently poorly understood.
It has been shown that the circulating concentration of juvenile hormone (JH) increases during the lifespan of the honeybee, and that the levels of JH correspond to age-related tasks. In addition, it has been shown that JH lowers the behavioral threshold to many task-related stimuli (Robinson, 1987), suggesting that JH may play a role in controlling the onset of age-related task performance.
Furthermore, young bees raised in isolation show increased levels of JH and the tendency to engage in foraging activity (Huang and Robinson, 1992). These data suggest that the presence of the older bees in a nest may inhibit the production of JH in young bees, thus preventing them from engaging in activities normally performed by older bees. According to the model, as the older bees die, younger bees receive less inhibition and thus have increased levels of JH, promoting behavioral patterns appropriate for older bees.
In this model, the processing of task-related olfactory stimuli (i.e. alarm pheromones) changes in response to JH. The possible neural substrates of this processing are shown below:
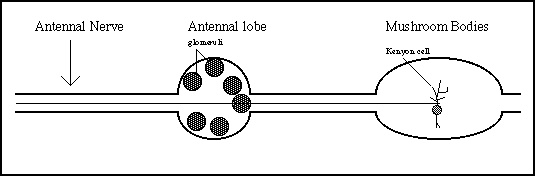
Schematic diagram of the honeybee olfactory system
Inputs from the antennal nerve project to the glomeruli of the antennal lobe. There a two categories of AL neurons: local interneurons and projection neurons. The specific information-processing role of the interneurons is not known, but recent data indicates that they are GABAergic, and hence inhibitory. The output neurons project through the antenno-glomerular tracts to the Kenyon cells of the mushroom bodies. The mushroom bodies also receive visual input and have been postulated to have a role in learning and memory (Menzel and Muller, 1996).
Data from Robinson (1987) indicates that the changes in olfactory processing of alarm pheromones are not mediated by the antennal chemoreceptors. This implicates the central olfactory processing system (AL and mushroom bodies) in mediating this plasticity and hence age-based division of labor. Future neurobiological investigation should help elucidate the neural mechanisms involved in this plasticity and will help us to understand some of the principles by which this highly complex social network of bees is organized.
Good Overview of Honeybee Neurobiology:
Menzel R. and Mercer A. eds (1987) Neurobiology and Behavior of Honeybees.
Springer-Verlag.
Activator-Inhibitor Model of Division of Labor:
Huang Z-Y and G. Robinson (1982) Honeybee colony integration: Worker- worker
interactions mediate hormonally regulated plasticity in division of labor.
Proc. Natl. Acad. Sci. 89: 11726-11729.
Robinson G. (1987) Modulation of alarm pheromone perception in the honey bee: Evidence for division of labor based on hormonally regulated response thresholds. J. Comp. Physiol. A. 160: 613-619.
Anatomy and Physiology of Honeybee Olfactory System:
Flanagan D. and A. Mercer (1989) Morphology and response characteristics
of neurons in the deutocerebrum of the honeybee Apis mellifera. J. Comp.
Physiol. A 164: 483-494.
Iwama A., Sugihara D. and T. Shibuya (1995) Morphology and physiology of neurons responding to the Nasonov pheromone in the antennal lobe of the honeybee, Apis mellifera. Zoological Science. 12: 207-218.
Plasticity in Honeybee Nervous System:
Menzel R. and U. Muller (1996) Learning and memory in honeybees: From behavior
to neural substrates. Ann. Rev. Neurosci. 19: 379-404.
