
Mustached Bat, Pteronotus parnellii
Bats that echolocate are of the suborder Microchiroptera within the mammalian order Chiroptera. Within the Microchiropteran suborder there are over 800 species of bats, and these bats produce all manner of sounds for echolocation, from bisonar pulses to clicks and other calls. Many of the Microchiropteran bats that echolocate do so for the purpose of catching prey, mainly insects. Therefore their echolocation abilities are a premier sensory modality in their nervous systems. Bats like the mustached bat pictured above have nervous systems highly dedicated to the task of echolocation, and are an excellent species to study from a neuroethological perspective.
Tree of Life: Chiroptera
Interestingly enough, the physical movement of bats like the mustached
bat can be observed, but their echolocation abilities cannot be heard by
the human ear. The bat's sonar pulses are well beyond the range of human
hearing; bats produce sounds of 30KHz and higher! However, a great deal
is known about the types of sounds that the echolocating bats produce, and
the types of sounds that they hear in return, thanks to the large body of
literature that exists on echolocation.
In order to find its prey the bat may sit on a perch or fly around using
its sonar signals as information about food in the area. The mustached bat
is considered a 'high duty cycle' bat since it produces signals 80% of the
time that it spends echolocating. The sonar signals that the bat produces
return to it in the form of an echo, which the bat can detect with its auditory
structures. The echo that is reflected back by the target carries information
about the size of the target based on the energy reflected from the pulse.
Bats do not all produce the same types of sounds or calls. One of the most
highly studied bats, the mustached bat, produces a bisonar sound consisting
of a constant frequency portion (CF) followed by a downward frequency modulated
sweep (FM), often expressed by the sound 'iiiiiiiiiu'. This has lead to
the mustached bat being classified in the literature as a CF-FM bat, along
with a few other species. Other Microchiropteran bats produce only the CF
component or only the FM components. It turns out that the CF and FM portions
of the sonar pulse serve different purposes.
The constant frequency portion of a pulse is great for detecting targets
and measuring the Doppler shift. The FM portion of a pulse is excellent
for honing in on the distance of an object and some of its finer details.
This choice between CF and FM may also have to do with the conditions in
which the bat hunts. The mustached bat often hunts in vegetation and the
CF component of the pulse may help this bat focus in on insects moving within
the vegetation.
When the mustached bat produces a CF-FM pulse, it also produces harmonics
of that pulse. Thus the bat is not only getting info from its first frequency,
but also the 2nd, 3rd, and 4th frequencies of that pulse.
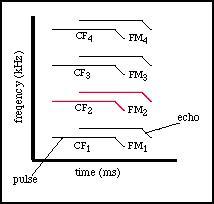
Of particular interest to the mustached bat are the echoes returning
from its 2nd harmonic pulse. This echo returns to the bat with a great deal
of information about the Doppler shift of the target. And there is a large
specialization in the auditory cortex of the bat that focuses on the frequency
range of this echo.
When a bat begins to echolocate it usually produces short millisecond long
pulses of sonar (CF-FM in the case of the mustached bat), and listens to
the returning echoes. If prey is detected by the bat, it will generally
fly toward the source of the echo continuing to emit sounds and focus in
more accurrately on the prey. As the bat gets closer and closer to the target
the sonar pulses are emitted faster with a shorter duration. This happens
until the bat is right upon the prey, at which time the bat scoops the insect
up into its wing membranes and into its awaiting mouth. It's a beautiful,
complicated and highly accurate form of prey capture that is thoroughly
supported by the bat's innate neural mechanisms.
It is important to remember that although bats are creatures of flight, they are mammals. With that distinction can come some very complicated biology, especially within the sensory modalities. Many researchers have dedicated their entire careers to studying how it is that bats echolocate with such great precision.
Bats have auditory systems that are like many other auditory systems in mammals. They hear sounds through their ears which directs the sound through the inner ear and onto the basilar membrane of the cochlea. The basilar membrane in turn vibrates according to the frequency of the sound and turns that mechanical signal into a neural code that is carried into the brainstem and to the rest of the brain. Even at the level of the basilar membrane it becomes obvious just how dedicated the auditory system is to the task of echolocation. The basilar membrane itself in the mustached bat, Pteronotus parnellii is thickened precisely at the frequencies that the bat is most interested in, 61.0-61.5 KHz. These frequencies correspond to the returning echoes of the Doppler-shift compensated 2nd harmonic of the CF-FM pulse.
This specialization for frequencies between 61.0-61.5 KHz will be observed throughout the auditory system of the mustached bat. Individual spiral ganglion cells of the bat's brain also show a sharp tuning to one particular frequency in this range. And the whole range is well represented with many spiral ganglion cells. According to Nobuo Suga, a key researcher in this field, the mustached bat has developed this specialization for these frequencies because of the conditions that it hunts in. The mustached bat often hunts in dense vegetation, which would normally add a lot of background 'noise' to the bat's hunting if it were just using an FM signal. However, the mustached bat uses a CF-FM pulse to detect its prey, especially focusing on the 2nd harmonic of the CF portion of the pulse to give it specific information. The basilar membrane specializations allow the bat to be sensitive to the returning echo, but not as sensitive to the emitted pulse. And the sharp tuning curves of the spiral ganglion cells equip the bat with a way to cut down on background noise from the surroundings. Since the cells are so sharply tuned, they will not be excited by just any frequencies returning from the periphery, but instead will be excited by the 'right' frequency from a potential meal.
There are so many pieces of information coded into the pulses and echoes that a bat hears, it is impossible to cover them all in one summary. The following is a list of cues that the bat can discern from what it hears: relative velocity of a target, flutter of target, range, size, fine characteristics, azimuth and elevation. This information is encoded in the amplitude of returning echoes, in the delay between echo and pulse, the Doppler shift between pulse and echo, and many other combinations of factors. The bat appears to be an amazing signal processing machine that has an accuracy of 99%!
This accuracy does not come from specializations at the auditory periphery
alone, many other higher brain structures in the bat are specialized for
this analysis as well. Three of the most amazing regions of analytical specialization
occur at the level of the auditory cortex: the CF-CF area (dark orange),
the FM-FM area (light green), and the DSCF area (pink).
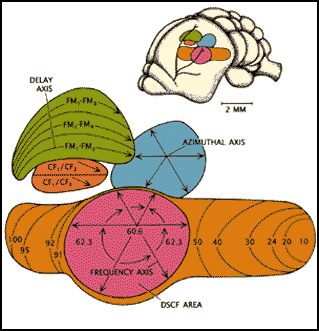
A schematic representation of bat auditory cortex
For some time the importance of the Doppler shift has been eluded to in this summary. The Doppler shift is a phenomena that occurs with sound waves. As a bat produces pulses of sonar they are reflected back from a target. However, the pulses are not just reflected back 'as is,' there is some compression of the sound waves which causes them to reflect back at a higher frequency. In the case of an insect, the wing beat of the insect causes its own Doppler shift to be superimposed on the existing Doppler shift. The mustached bat can detect these ripples in the Doppler shift and this leads them to their insect target.
The mustached bat engages in what is known as Doppler-shift compensation,
that is the bat lowers its own emitted CF so that the Doppler shifted echo
returning from the insect is precisely in the range where the bat has the
best chance of detecting it...61.0-61.5 KHz. Thus the specialization at
the level of the basilar membrane and spiral ganglion makes intuitive sense.
But the bat takes this analysis one level higher. In the tonotopic organization
of the auditory cortex there is a relatively large portion of the cortex
dedicated to the frequencies between 60.6 and 62.3 KHz. This is known as
the DSCF area (Doppler shifted constant frequency area), illustrated in
pink in the above figure. This area's frequencies are specialized for the
mustached bat based on its resting frequency of 61 KHz; however, another
species of bat with a different resting frequency would have its own frequency
focus. This area is often referred to as an 'acoustic fovea', drawing a
comparison between the fovea in the visual system of some animals and the
DSCF area in bats.
The neurons of the DSCF area have a particular frequency and amplitude that
will excite them maximally. The cells are also arranged in a columnar fashion,
similar to the cytoarchitecture in the visual cortex of the cat. Following
that analogy, cells in perpendicular colums in the DSCF area respond to
one particular frequency and amplitude. Unlike other areas in the bat's
cortex, cells in the DSCF area only respond to the frequency and amplitude
of the echo of the 2nd harmonic of the CF. They do not respond to the emitted
pulse. The area is hypothesized to be used in discriminating minute differences
in frequency, the sort that would cause a flying insect to appear above
the background noise and existing Doppler shift. Suga has hypothesized that
this is the "region responsible for the precision of the Doppler-shift
compensation but not for performing the actual compensation." It is
still a mystery as to how the DSCF area connects to the structures responsible
for that important task.
Another area of interest that also relates to the bat's use of Doppler shift information is the CF/CF area of auditory cortex, the small dark orange region in the figure above. This is the area in the cortex where the bat calculates the actual Doppler-shift from the target. The bat does this by comparing the frequencies between the CF pulse and its 2nd or 3rd harmonic echo. Thus there are two classes of CF/CF neurons, each for the corresponding pulse echo pairs. These neurons in the CF/CF area respond maximally to stimulation from the produced pulse and returning echo, this is what classifies them as a mechanism for neural integration at the cortical level.
As with the DSCF area, there is a particular architecture to the CF/CF
area. The cells are arranged according to the 1st harmonic of the CF longitudinally.
Along the width of the area, cells are arranged according to the frequency
of the returning echo. Therefore, at intersecting points in the CF/CF area
a functional map is created that corresponds to the specific relative target
velocity, and this ranges from -2 to 9 meters per second. It has been found
that the velocities from zero to 4 meters per second are overrepresented
in that map because of the bats need for precision at those speeds for landing
or catching prey.
The steps to the CF/CF area, DSCF area and FM/FM (which there is not sufficient
room to discuss) are diagrammed in the figure below. The colors in the figure
correspond to the colors in the first figure in the neural substrates section.
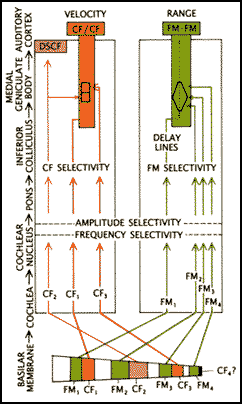
Gross connectivity from auditory input to cortex
Although there is more to echolocation in bats than what has been mentioned in this web page, I believe it is clear how exquisite the system for the production and analysis of echolocation signals is in the bat. Many structures at all levels of the nervous system, from basilar membrane to cortex, have been dedicated to permitting the mustached bat to navigate and catch prey. As research continues on this species and others, more information is being gained on what occurs at the level of the auditory brainstem and the thalamus that permits for such detailed processing at the cortical level. It seems impossible to approach the bat from anything but a neuroethological perspective, since it appears that each specialization is driven by the environment of the bat, and the particular frequencies that it produces.
All pictures and figures from Suga, N. (1990). Bisonar
and neural computation in bats. Sci. Amer. 262:60-68.
Visit these other 'batty' sites for more information !
Jim Buzbee's bat house site has every bat link imaginable...check it out!
Bat Conservation International's web site
The University of Michigan's collection of information on bats
An amazing archive
of bat pictures!
Griffin, D.R., Webster F.A., Michael, C.R. (1960). The echolocation of flying insects by bats. Animal Behav. 8:151-154.
Schnitzler, H.U., Flieger, E. (1983). Detection of oscillating target movements by echolocation in the greater horshoe bat. J. Comp Physiol. 153:385-391.
Simmons, J.A. (1979). Perception of echo phase in bat sonar. Science 204:1336-1338.
Suga, N., Simmons, J.A., Jen, P.S. (1975). Peripheral specialization for fine analysis of Doppler-shifted echoes in the auditory system of the CF-FM bat Pteronotus parnellii. J Exp Biol. 63:161-192.
Fenton, M.B. (1995). Natural history and bisonar signals. In Hearing
by Bats.
Popper, A.N. Fay, R.R. (Eds). pp. 37-86. Springer Verlag, NY.
Grinnell, A.D. (1995). Hearing in bats: an overview. In Hearing by Bats. Popper, A.N. Fay, R.R. (Eds). pp. 1-36. Springer Verlag, NY.
Simmons, J.A., Howell, D.J., Suga, N. (1975). Information content of bat sonar echoes. Amer. Scientist. 63:204-215.
Suga, N. (1990) Bisonar and neural computation in bats. Sci. Amer. 262:60-68.
Dror, I.E., Zagaeski, M., Moss, C.F. (1995). Three-dimensional target recognition via sonar: a neural network model. Neural Networks. 8:149 160.
Kuc, R. (1994). Sensorimotor model of bat echolocation and prey capture. J. Acoust. Soc. Amer. 96:1965-1978.
Vater, M., Casseday, J.H., Covey, E. (1995). Convergence and divergence of ascending binaural and monaural pathways from the superior olives of the mustached bat. J. Comp. Neurol. 351:632-646.
